Complementary Interactions between Proteins and Ligands: The Immune System and Immunoglobulins: -Self Is Distinguished from Nonself by the Display of Peptides on Cell Surfaces
The immune system must identify and destroy pathogens, but it must also recognize and not destroy the normal proteins and cells of the host organism—the “self.” Detection of protein antigens in the host is mediated by MHC (major histocompatibility complex) proteins. MHC proteins bind peptide fragments of proteins digested in the cell and present them on the outside surface of the cell. These peptides normally come from the digestion of typical cellular proteins, but during a viral infection viral proteins are also digested and presented on the cell surface by MHC proteins. Peptide fragments from foreign proteins that are displayed by MHC proteins are the antigens the immune system recognizes as nonself. T-cell receptors bind these fragments and launch the subsequent steps of the immune response. There are two classes of MHC proteins (Fig. 5–21), which differ in their distribution among cell types and in the source of digested proteins whose peptides they display.
Class I MHC proteins (Fig. 5–22) are found on the surface of virtually all vertebrate cells. There are countless variants in the human population, placing them among the most polymorphic of proteins. Because each individual produces up to six class I MHC protein variants, any two individuals are unlikely to have the same set. Class I MHC proteins bind and display peptides de rived from the proteolytic degradation and turnover of proteins that occurs randomly within the cell. These complexes of peptides and class I MHC proteins are the recognition targets of the T-cell receptors of the TC cells in the cellular immune system. The general pattern of immune system recognition was first described by Rolf Zinkernagel and Peter Doherty in 1974. Each TC cell has many copies of only one T-cell receptor that is specific for a particular class I MHC protein–peptide complex. To avoid creating a legion of TC cells that would set upon and destroy normal cells, the maturation of TC cells in the thymus includes a stringent selection process that eliminates more than 95% of the developing TC cells, including those that might recognize and bind class I MHC proteins displaying pep tides from cellular proteins of the organism itself. The TC cells that survive and mature are those with T-cell receptors that do not bind to the organism’s own proteins. The result is a population of cells that bind foreign peptides bound to class I MHC proteins of the host cell. These binding interactions lead to the destruction of parasites and virus-infected cells. Following organ transplantation, the donor’s class I MHC proteins, recognized as foreign, are bound by the recipient’s TC cells, leading to tissue rejection. Class II MHC proteins occur on the surfaces of a few types of specialized cells, including macrophages and B lymphocytes that take up foreign antigens. Like class I MHC proteins, the class II proteins are highly polymorphic, with many variants in the human population. Each human is capable of producing up to 12 variants, and thus it is unlikely that any two individuals have an identical set. The class II MHC proteins bind and dis play peptides derived not from cellular proteins but from external proteins ingested by the cells. The resulting class II MHC protein–peptide complexes are the binding targets of the T-cell receptors of the various helper T cells. TH cells, like TC cells, undergo a stringent selection process in the thymus, eliminating those that recognize the individual’s own cellular proteins.
Despite the elimination of most TC and TH cells dur ing the selection process in the thymus, a very large number survive, and these provide the immune response. Each survivor has a single type of T-cell receptor that can bind to one particular chemical structure. The T cells patrolling the bloodstream and the tissues carry millions of different binding specificities in the T cell receptors. Within the highly varied T-cell population there is almost always a contingent of cells that can specifically bind any antigen that might appear. The vast majority of these cells never encounter a foreign anti gen to which they can bind, and they typically die within a few days, replaced by new generations of T cells end lessly patrolling in search of the interaction that will launch the full immune response. The TH cells participate only indirectly in the de struction of infected cells and pathogens, stimulating the selective proliferation of those TC and B cells that can bind to a particular antigen. This process, called clonal selection, increases the number of immune system cells that can respond to a particular pathogen. The importance of TH cells is dramatically illustrated by the epidemic produced by HIV (human immunodeficiency virus), the virus that causes AIDS (acquired immune deficiency syndrome). The primary targets of HIV infection are TH cells. Elimination of these cells progressively incapacitates the entire immune system.
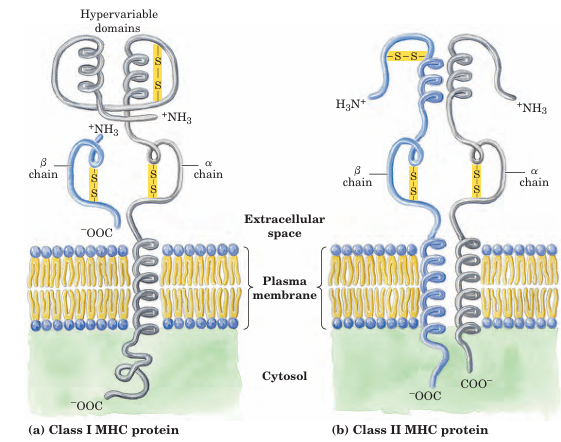
FIGURE 5–21 MHC proteins. These proteins consist α of β and chains. (a) In class I MHC proteins, the small β chain is invariant but the amino acid sequence of the α chain exhibits a high degree of variability, localized in specific domains of the protein that appear on the outside of the cell. Each human produces up to six different α chains for class I MHC proteins. (b) In class II MHC proteins, both the α and β chains have regions of relatively high variability near their amino-terminal ends.
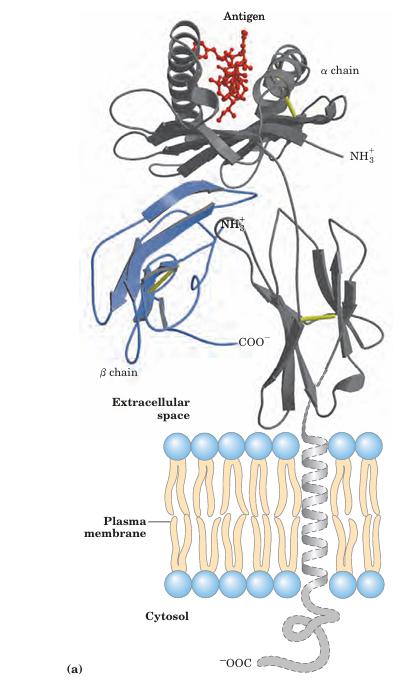
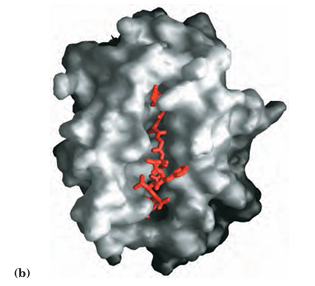
FIGURE 5–22 Structure of a human class I MHC protein. (a) This model is derived in part from the known structure of the extracellular portion of the protein (PDB ID 1DDH). The chain of MHC is shown in gray; the small β chain is blue; the disulfide bonds are yellow. A bound ligand, a peptide derived from HIV, is shown in red. (b) Top view of the protein, showing a surface contour image of the site where peptides are bound and displayed. The HIV peptide (red) occupies the site. This part of the class I MHC protein interacts with T-cell receptors.





















