























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Metabolism Regulation
 المؤلف:
Denise R. Ferrier
المؤلف:
Denise R. Ferrier
 المصدر:
Lippincott Illustrated Reviews: Biochemistry
المصدر:
Lippincott Illustrated Reviews: Biochemistry
 الجزء والصفحة:
الجزء والصفحة:
 16-9-2021
16-9-2021
 2752
2752




+
-
20
Metabolism Regulation
The pathways of metabolism must be coordinated so that the production of energy or the synthesis of end products meets the needs of the cell. Furthermore, individual cells function as part of a community of interacting tissues, not in isolation. Thus, a sophisticated communication system has evolved to coordinate the functions of the body. Regulatory signals that inform an individual cell of the metabolic state of the body as a whole include hormones, neurotransmitters, and the availability of nutrients. These, in turn, influence signals generated within the cell (Fig. 1).
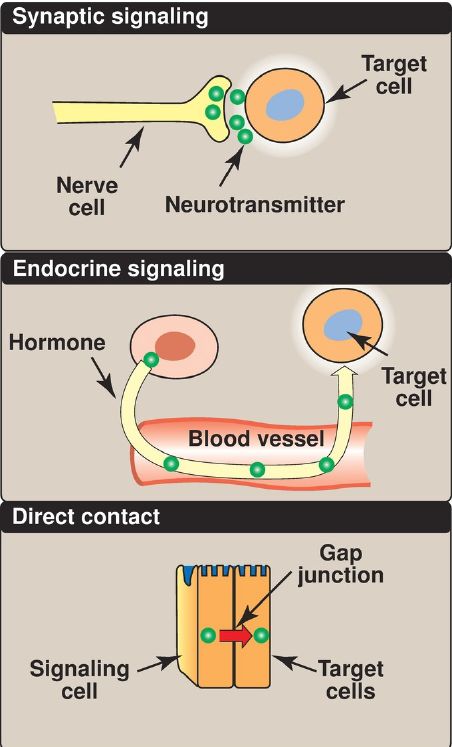
Figure 1: Some commonly used mechanisms for transmission of regulatory signals between cells.
A. Intracellular communication
The rate of a metabolic pathway can respond to regulatory signals that arise from within the cell. For example, the rate may be influenced by the availability of substrates, product inhibition, or alterations in the levels of allosteric activators or inhibitors. These intracellular signals typically elicit rapid responses and are important for the moment-to-moment regulation of metabolism.
B. Intercellular communication
The ability to respond to intercellular signals is essential for the development and survival of organisms. Signaling between cells provides for long-range integration of metabolism and usually results in a response, such as a change in gene expression, that is slower than is seen with intracellular signals. Communication between cells can be mediated, for example, by surface-to-surface contact and, in some tissues, by formation of gap junctions, allowing direct communication between the cytoplasms of adjacent cells. However, for energy metabolism, the most important route of communication is chemical signaling between cells by blood-borne hormones or by neurotransmitters.
C. Second messenger systems
Hormones and neurotransmitters can be thought of as signals and their receptors as signal detectors. Receptors respond to a bound ligand by initiating a series of reactions that ultimately result in specific intracellular responses. Second messenger molecules, so named because they intervene between the original extracellular messenger (the neurotransmitter or hormone) and the ultimate intracellular effect, are part of the cascade of events that converts (transduces) ligand binding into a response. Two of the most widely recognized second messenger systems are the calcium/phosphatidylinositol system and the adenylyl cyclase (adenylate cyclase) system, which is particularly important in regulating the pathways of intermediary metabolism. Both involve the binding of ligands, such as epinephrine or glucagon, to specific G protein–coupled receptors (GPCR) on the cell (plasma) membrane. GPCR are characterized by an extracellular ligand-binding domain, seven transmembrane α helices, and an intracellular domain that interacts with trimeric G proteins (Fig. 2). [Note: Insulin, another key regulator of metabolism, binds a membrane tyrosine kinase receptor and not a GPCR.]
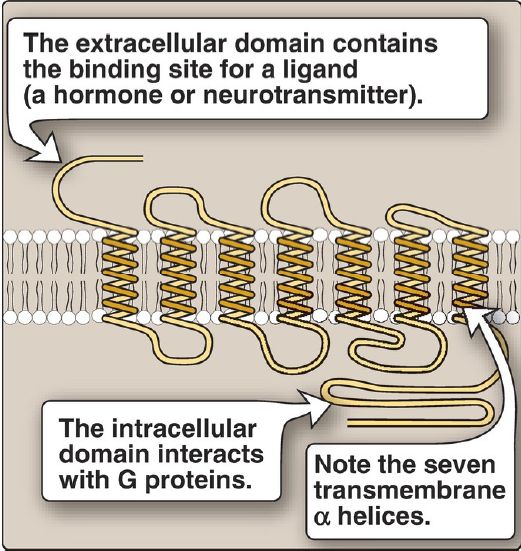
Figure 2: Structure of a typical G protein–coupled receptor of the plasma membrane.
D. Adenylyl cyclase
The recognition of a chemical signal by some GPCR, such as the β- and α2-adrenergic receptors, triggers either an increase or a decrease in the activity of adenylyl cyclase (AC). This is a membrane-bound enzyme that converts ATP to 3ʹ,5ʹ-adenosine monophosphate (cyclic AMP, or cAMP). The chemical signals are most often hormones or neurotransmitters, each of which binds to a unique type of GPCR. Therefore, tissues that respond to more than one signal must have several different GPCR, each of which can be linked to AC.
1. Guanosine triphosphate–dependent regulatory proteins: The effect of the activated, occupied GPCR on second messenger formation is indirect, mediated by specialized trimeric proteins (α, β, and γ subunits) of the cell membrane. These proteins, referred to as G proteins because the α subunit binds guanosine di- or triphosphates (GDP or GTP), form a link in the chain of communication between the receptor and AC. In the inactive form of a G protein, the α subunit is bound to GDP (Fig. 3).
Ligand binding causes a conformational change in the receptor, triggering replacement of this GDP with GTP. The GTP-bound form of the α subunit dissociates from the βγ subunits and moves to AC, affecting enzyme activity. Many molecules of active Gα protein are formed by one activated receptor. [Note: The ability of a hormone or neurotransmitter to stimulate or inhibit AC depends on the type of Gα protein that is linked to the receptor. One type, designated Gs, stimulates AC (see Fig. 3), whereas another type, designated Gi, inhibits the enzyme (not shown).]
The actions of the Gα–GTP complex are short-lived because Gα has an inherent GTPase activity, resulting in the rapid hydrolysis of GTP to GDP. This causes inactivation of Gα, its dissociation from AC, and its reassociation with the βγ dimer.
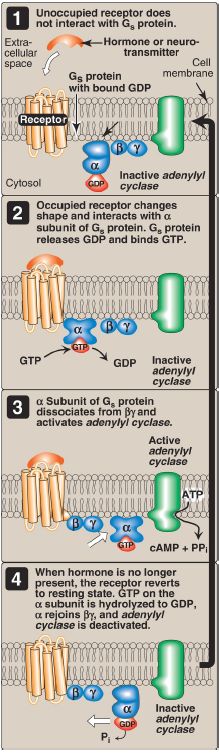
Figure 3: The recognition of chemical signals by certain membrane receptors triggers an increase (or, less often, a decrease) in the activity of adenylyl cyclase. GDP and GTP = guanosine di- and triphosphates; cAMP = cyclic adenosine monophosphate.
Toxins from Vibrio cholerae (cholera) and Bordetella pertussis (whooping cough) cause inappropriate activation of AC through covalent modification (ADP-ribosylation) of different G proteins. With cholera, the GTPase activity of Gαs is inhibited in intestinal cells. With whooping cough, Gαi is inactivated in respiratory tract cells.
2. Protein kinases: The next step in the cAMP second messenger system is the activation of a family of enzymes called cAMP-dependent protein kinases such as protein kinase A (PKA), as shown in Figure 4. cAMP activates PKA by binding to its two regulatory subunits, causing the release of its two catalytically active subunits. These subunits transfer phosphate from ATP to specific serine or threonine residues of protein substrates. The phosphorylated proteins may act directly on the cell’s ion channels or, if enzymes, may become activated or inhibited. [Note: Several types of protein kinases are not cAMP dependent, for example, protein kinase C]
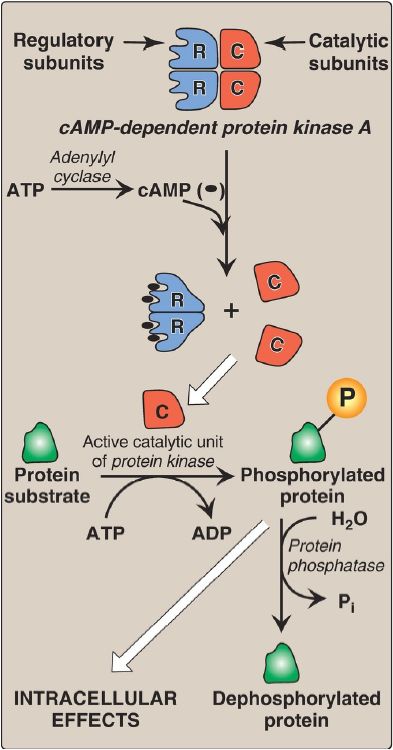
Figure 5: Actions of cyclic adenosine monophosphate (cAMP). = phosphate; ADP = adenosine diphosphate; Pi = inorganic phosphate.
3. Protein phosphatases: The phosphate groups added to proteins by protein kinases are removed by protein phosphatases, enzymes that hydrolytically cleave phosphate esters (see Fig. 5). This insures that changes in protein activity induced by phosphorylation are not permanent.
4. cAMP hydrolysis: cAMP is rapidly hydrolyzed to 5ʹ-AMP by cAMP phosphodiesterase that cleaves the cyclic 3ʹ,5ʹ-phosphodiester bond. 5ʹ- AMP is not an intracellular signaling molecule. Therefore, the effects of neurotransmitter- or hormone-mediated increases of cAMP are rapidly terminated if the extracellular signal is removed. [Note: cAMP phosphodiesterase is inhibited by caffeine, a methylxanthine derivative.]
 الاكثر قراءة في الكيمياء الحيوية
الاكثر قراءة في الكيمياء الحيوية
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















